Become an IES Member today
Join our community of lighting professionals and enjoy the benefits

Register Now or Become a sponsor
Join us for the 2024 IES & DOE Research Symposium Learn and connect at this exclusive event

IES Illuminance Selector
Design your projects to IES/ANSI recommended light levels faster than ever though the Illuminance Selector

Become a Conference Partner
Sponsor and exhibit bookings are now open for IES24, The Lighting Conference Don’t miss this opportunity to network with over 500 professionals at the State of the Industry Event

Become a Sustaining Member
IES Sustaining Membership is for companies, institutions, and other organizations regardless of staff size or operational budget.

Featured Event
International Day of Light Research Webinar: Focus on Roadways and Safety
Join us for a panel focusing on some of the research being done to improve safety on the road and in the field. The projects being showcased include adaptive roadway…
IES & DOE 2024 Research Symposium
New York City, August 15-17, New York Marriott Marquis, Times Square Symposium dates: 15 – 16 May, 2024 Location: OMSI, 1945 SE Water Ave, Portland, OR 97214 New this year!…
International Day of Light Research Webinar: Focus on Daylighting
The authors of three papers recently published in Leukos will present their work on various aspects of daylight and daylighting. The projects being showcased include: Sunlight Autonomy for Buildings: A New…
International Day of Light Research Webinar: Focus on Roadways and Safety
The IES is on a mission to improve life through quality of light.
Wherever you are in your professional journey, we have the resources to support you.
The IES is on a mission to improve life through quality of light.
Wherever you are in your professional journey, we have the resources to support you.
A Third-Place Finish

How They Did It: Tiffany Window
To illuminate Light in Heaven and Earth with electric lights for The Art Institute of Chicago, designers from Schuler Shook worked closely with museum curators, conservators and exhibit designers to…

Spectral Light Meter Obtains WELL Eligibility
The MSC15-CS spectral circadian light meter by Germany-based manufacturer Gigahertz-Optik, known for UV-VIS-NIR optical radiation measurement tools, has received the Works with WELL trademark offered through the International WELL Building…

LIA, Loughborough University, Holophane Europe Join Forces for Students
In an ongoing effort to encourage the next generation of students to find successful career opportunities in the lighting design industry, the Lighting Industry Association (LIA) and Holophane Europe, Ltd.—a…
LIGHTING DESIGN & APPLICATION

Jet-Setters: Designers Discuss the Adventure of International Projects

Message: We Care
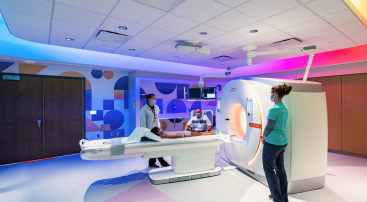
Healthy Choices: A Medical Center Leverages Lighting to Let Kids be Kids

A Third-Place Finish
How They Did It: Tiffany Window
Spectral Light Meter Obtains WELL Eligibility
Join Our Mailing List
Thank you for generously supporting our efforts to improve life through quality of light.





